Для максимально полного изучения балто-славянских популяций генетики использовали данные трёх типов: Y-хромосома (6078 образцов из 62 популяций), мтДНК (6876 образцов из 48 популяций), и широкогеномные (полногеномные) маркеры (1 297 образцов из 16 популяций). Их анализ позволяет группировать геномы в классы с общими SNP (гаплогруппы). Выявленные точечные мутации позволили установить время расхождения разных генетических линий их носителей.
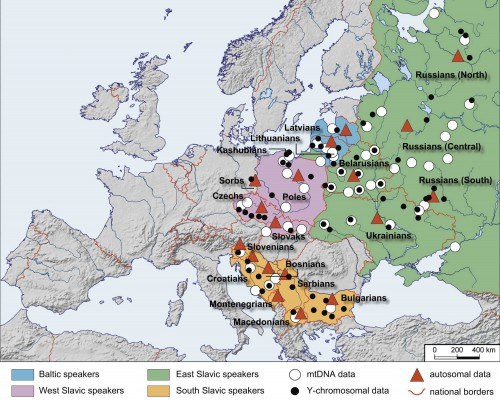 По трём генетическим системам были изучены народы, говорящие на языках балто-славянской лингвистической группы – шестнадцать народов по единой обширной панели маркеров:
балтские народы – латыши и литовцы;
восточные славяне – белорусы, русские, украинцы и русины;
западные славяне – кашубы, поляки, словаки, сорбы и чехи;
южные славяне – болгары, боснийцы, македонцы, сербы, словенцы и хорваты.
По трём генетическим системам были изучены народы, говорящие на языках балто-славянской лингвистической группы – шестнадцать народов по единой обширной панели маркеров:
балтские народы – латыши и литовцы;
восточные славяне – белорусы, русские, украинцы и русины;
западные славяне – кашубы, поляки, словаки, сорбы и чехи;
южные славяне – болгары, боснийцы, македонцы, сербы, словенцы и хорваты.
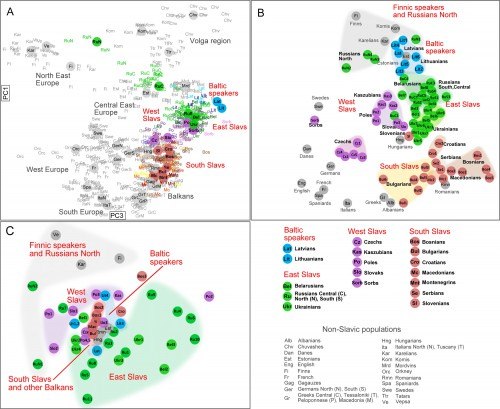 Восточные славяне – русские, белорусы, украинцы и русины – отчётливо группируются в единый генетический кластер. Исключение составляют русскоязычные популяции северного ареала, которые генетически отдалены от восточных славян и тяготеют к соседним финно-угорским и скандинавским популяциям.
Из западных славян: чехи, и в меньшей степени словаки, отличаются от восточных славян, и смещены в сторону германоязычных народов и других западноевропейских популяций. В то время как поляки наиболее близки к восточным славянам.
Южные славяне, формируют дисперсную группу, которая внутренне поделена на: западные (словенцы, хорваты и боснийцы) и восточные (македонцы и болгары) регионы, с сербами посередине. При этом, словенцы генетически близки к венграм, а восточная ветвь южных славян группируется с румынами и до некоторой степени с греками.
Балтские народы – латыши и литовцы – имеют генетическую близость к эстонцам и к западным беларусам. Оказалось также, что балтские популяции близки к волжской группе финно-угорских народов (особенно к эрзянам).
Восточные славяне – русские, белорусы, украинцы и русины – отчётливо группируются в единый генетический кластер. Исключение составляют русскоязычные популяции северного ареала, которые генетически отдалены от восточных славян и тяготеют к соседним финно-угорским и скандинавским популяциям.
Из западных славян: чехи, и в меньшей степени словаки, отличаются от восточных славян, и смещены в сторону германоязычных народов и других западноевропейских популяций. В то время как поляки наиболее близки к восточным славянам.
Южные славяне, формируют дисперсную группу, которая внутренне поделена на: западные (словенцы, хорваты и боснийцы) и восточные (македонцы и болгары) регионы, с сербами посередине. При этом, словенцы генетически близки к венграм, а восточная ветвь южных славян группируется с румынами и до некоторой степени с греками.
Балтские народы – латыши и литовцы – имеют генетическую близость к эстонцам и к западным беларусам. Оказалось также, что балтские популяции близки к волжской группе финно-угорских народов (особенно к эрзянам).
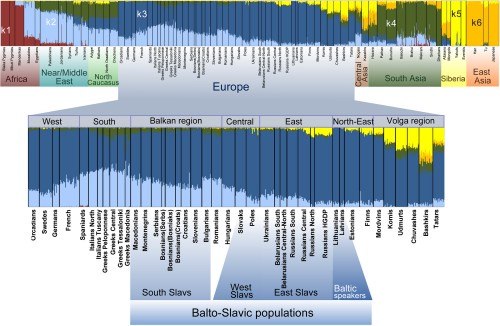 У балто-славянских популяций почти весь спектр представлен двумя основными цветами предковых компонентов: синий (k3) и голубой (k2). Предковый компонент k3 максимален у балтских популяций, превалирует у восточных славян (80-95%) и снижается у южных славян (55-70%). Напротив, k2 (голубой) более характерен для популяций средиземноморского и кавказского регионов и снижается к северу Европы. У южных славян на него приходится примерно 30%, у западных славян происходит снижение до 20%, а у северных русскоязычных и балтских популяций – до 5%.
У славянской этногруппы также присутствует лимонно-жёлтый компонент в предковом спектре, обозначаемый как k5, который значительно распространён только среди восточных славян, и в особенности среди северного кластера. По происхождению этот компонент является сибирским и составляет основную часть спектра для популяций Сибири. Тёмно-зелёный компонент k4 характерен для популяций Южной Азии, а также распространен на Ближнем Востоке и Средиземноморье и встречается низкой частотой у южных славян и других народов Балканского полуострова.
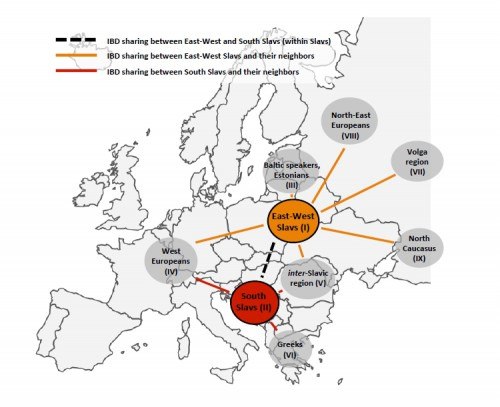
Популяционными генетиками было подсчитано число общих гаплотипов между «западно-восточными» славянами и восемью другими группами народов Европы:
1) южными славянами (болгары, боснийцы, македонцы, словенцы, хорваты);
2) популяциями Западной Европы (итальянцы, немцы, французы);
3) балтскими популяциями (латыши, литовцы);
4) популяциями северо-восточной Европы (вепсы, карелы, финны, эстонцы);
5) популяциями центральной Европы, ареал которых находится между западно-восточными и южными славянами – «между-славянские популяции»; разноязычные популяции: гагаузы, венгры, румыны);
6) греками;
7) популяциями волжского региона и Приуралья (башкиры, коми, эрзя, мокша, татары, удмурты, чуваши);
8) северокавказскими популяциями (адыги, балкарцы, ногайцы).
При введении условного «балто-славянского эталона» общих гаплотипов, часть окружающих неславянских популяций будет иметь меньшее родство с западно-восточными славянами, чем южные славяне – народы Поволжья, Западной Европы, Кавказа, а также греки. Родство генофондов балтов и популяций северо-восточной Европы в два раза выше (вепсы, карелы, финны, латыши, литовцы, поморы, эстонцы).
У балто-славянских популяций почти весь спектр представлен двумя основными цветами предковых компонентов: синий (k3) и голубой (k2). Предковый компонент k3 максимален у балтских популяций, превалирует у восточных славян (80-95%) и снижается у южных славян (55-70%). Напротив, k2 (голубой) более характерен для популяций средиземноморского и кавказского регионов и снижается к северу Европы. У южных славян на него приходится примерно 30%, у западных славян происходит снижение до 20%, а у северных русскоязычных и балтских популяций – до 5%.
У славянской этногруппы также присутствует лимонно-жёлтый компонент в предковом спектре, обозначаемый как k5, который значительно распространён только среди восточных славян, и в особенности среди северного кластера. По происхождению этот компонент является сибирским и составляет основную часть спектра для популяций Сибири. Тёмно-зелёный компонент k4 характерен для популяций Южной Азии, а также распространен на Ближнем Востоке и Средиземноморье и встречается низкой частотой у южных славян и других народов Балканского полуострова.
Популяционными генетиками было подсчитано число общих гаплотипов между «западно-восточными» славянами и восемью другими группами народов Европы:
1) южными славянами (болгары, боснийцы, македонцы, словенцы, хорваты);
2) популяциями Западной Европы (итальянцы, немцы, французы);
3) балтскими популяциями (латыши, литовцы);
4) популяциями северо-восточной Европы (вепсы, карелы, финны, эстонцы);
5) популяциями центральной Европы, ареал которых находится между западно-восточными и южными славянами – «между-славянские популяции»; разноязычные популяции: гагаузы, венгры, румыны);
6) греками;
7) популяциями волжского региона и Приуралья (башкиры, коми, эрзя, мокша, татары, удмурты, чуваши);
8) северокавказскими популяциями (адыги, балкарцы, ногайцы).
При введении условного «балто-славянского эталона» общих гаплотипов, часть окружающих неславянских популяций будет иметь меньшее родство с западно-восточными славянами, чем южные славяне – народы Поволжья, Западной Европы, Кавказа, а также греки. Родство генофондов балтов и популяций северо-восточной Европы в два раза выше (вепсы, карелы, финны, латыши, литовцы, поморы, эстонцы).
 Согласно принципу эквидистантности, если система популяций развивается без больших внешних влияний, то постепенное разделение популяций приводит к линейному накоплению как лингвистического, так и генетического разнообразия. В результате генетическое разнообразие примерно одинаково на всех уровнях, будь то между популяциями одного народа, между этносами одной ветви или между их усредненными характеристиками. Величины генетического разнообразия славяноязычного населения на разных иерархических уровнях резко различаются – 0.06, 0.03, 0.01. Это указывает на активное смешение с окружающими народами. Наибольшая изменчивость приходится на самый древний уровень, что выявляет взаимодействия на ранних этапах истории славяноязычных популяций.
Тесты Y-хромосом, мтДНК и аутосомных маркеров показали чрезвычайно высокую корреляцию между генетикой и географическим положением популяций (0,80-0,95). Очень высокая корреляция также обнаружена между генетикой и лингвистикой (0,74-0,78). Однако, при исключении географического фактора, частная корреляция с лингвистикой становится намного ниже (0,3 для мтДНК и 0,2 для остальных двух систем), в то время как для всех трёх генетических систем корреляция с географией при исключении фактора лингвистики остается высокой (0,5 для мтДНК и 0,8 для остальных двух систем). Это указывает на то, что основная связь обусловлена географическим фактором, а высокая связь с лингвистикой объясняется тем, что народы, говорящие на родственных языках, часто являются и географическими соседями.
Согласно принципу эквидистантности, если система популяций развивается без больших внешних влияний, то постепенное разделение популяций приводит к линейному накоплению как лингвистического, так и генетического разнообразия. В результате генетическое разнообразие примерно одинаково на всех уровнях, будь то между популяциями одного народа, между этносами одной ветви или между их усредненными характеристиками. Величины генетического разнообразия славяноязычного населения на разных иерархических уровнях резко различаются – 0.06, 0.03, 0.01. Это указывает на активное смешение с окружающими народами. Наибольшая изменчивость приходится на самый древний уровень, что выявляет взаимодействия на ранних этапах истории славяноязычных популяций.
Тесты Y-хромосом, мтДНК и аутосомных маркеров показали чрезвычайно высокую корреляцию между генетикой и географическим положением популяций (0,80-0,95). Очень высокая корреляция также обнаружена между генетикой и лингвистикой (0,74-0,78). Однако, при исключении географического фактора, частная корреляция с лингвистикой становится намного ниже (0,3 для мтДНК и 0,2 для остальных двух систем), в то время как для всех трёх генетических систем корреляция с географией при исключении фактора лингвистики остается высокой (0,5 для мтДНК и 0,8 для остальных двух систем). Это указывает на то, что основная связь обусловлена географическим фактором, а высокая связь с лингвистикой объясняется тем, что народы, говорящие на родственных языках, часто являются и географическими соседями.
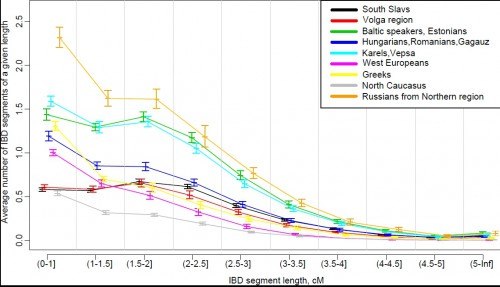 Генетики смогли выделить два основных субстрата, которые были ассимилированы славянами, впитав в себя местные популяции. «Центрально-восточноевропейский субстрат» был принят как западными, так и восточными славянами, с высокими частотами гаплогруппы R1a. Другой субстрат, «южно-восточноевропейский», был впитан южными славянами, с высокими частотами гаплогруппы I2a.
Генетики смогли выделить два основных субстрата, которые были ассимилированы славянами, впитав в себя местные популяции. «Центрально-восточноевропейский субстрат» был принят как западными, так и восточными славянами, с высокими частотами гаплогруппы R1a. Другой субстрат, «южно-восточноевропейский», был впитан южными славянами, с высокими частотами гаплогруппы I2a.
 Особенности Псковско-Новгородского ареала
Генетический анализ основного русскоязычного населения Восточно-Европейской равнины показывает, что основная часть этой популяции характеризуется низким уровнем генетической дифференциации как по мтДНК, так и по линиям Y-хромосомы. Генетические различия между популяциями исчезают, если их сгруппировать в соответствии с диалектным разнообразием русского языка и данными антропологии. Анализ популяций, представляющих этнографическую группу русскоязычных поморов, приводит к значительному увеличению значений межпопуляционной генетической дифференциации. Поморы отличаются высоким генетическим сходством с финно-угорским и, в некоторых случаях, со скандинавским населением Северной Европы, а также проявляют высокую степень многоступенчатости генофонда, вызванную влиянием генетического дрейфа.
Популяции объединяли в группы учитывая:
– (А) по говорам и наречиям русского языка: южное наречие (Белгород, Орёл, Калуга, Тула), северное наречие (Ярославль), среднерусские говоры (Великий Новгород, Волот, Псков, Владимир, Нижний Новгород);
– (Б) по диалектным зонам: юго-западная зона (Белгород, Орёл, Калуга, Тула), северо-западная зона (Великий Новгород, Волот, Псков), северо-восточная зона (Владимир, Нижний Новгород, Ярославль);
– (В) в соответствии с данными антропологии: предполагаемые потомки словен новгородских и псковских кривичей (Великий Новгород, Волот, Псков), кривичей (Владимир, Нижний Новгород, Ярославль), вятичей (Калуга, Тула), северян (Орёл, Белгород).
Анализ изменчивости нуклеотидных последовательностей гипервариабельного сегмента 1 (ГВС1) мтДНК в популяциях Тульской, Калужской, Владимирской, Ярославской, Псковской, Новгородской (Великий Новгород и Волот), Белгородской, Нижегородской, Орловской, Саратовской, Курской и Костромской областей и Ставропольского края (проанализировано 913 человек) – значение межпопуляционной дифференциации FST составляет 0,35% (P = 0,0007).
Анализ данных об изменчивости нуклеотидных последовательностей ГВС1 мтДНК в популяциях северо-западной зоны (Великий Новгород, Волот и Псков) был проведён в сравнении с другими славянскими популяциями Восточно-Европейской равнины (русские, поляки), а также популяциями соседних народов: балтскими (литовцы) и финскими (эстонцы, карелы и финны). Исследование показало, что степень дифференциации между проанализированными популяциями низка – FST = 0,38%, P = 0. Однако некоторые популяции демонстрируют высокое генетическое сходство.
Выделена отдельная группа населения, которая включает русскоязычные популяции северо-западной зоны (Великий Новгород и Псков), северо-восточную польскоязычную группу (Suwałki) и популяции балтийского и финского происхождения, что приводит к увеличению степени межгрупповой дифференциации (0,15%). Это свидетельствует о своеобразии генетической структуры этих популяций как по отношению к остальным славянским популяциям (русским и полякам; 0,52%), так и к популяциям балтийского и финского происхождения (0,76%). Анализ генофондов северо-западных русскоязычных и северо-восточных польскоязычных населений в сравнении с их непосредственными соседями – литовцами и эстонцами – показал значительные различия между ними (0,73%).
Таким образом, биологическое разнообразие является результатом формирования уникальной генетической композиции, сложившейся в результате взаимодействия трёх генофондов – славянского, балтского и финно-угорского. Анализ структуры митохондриального генофонда популяций Великого Новгорода, Пскова и Suwałki показал наличие только одной генетической компоненты – гаплогруппы U5a, которая распространена в этих популяциях с более высокой частотой (в среднем 16%), чем в соседних славянских, балтийских и финно-угорских популяциях, где её частота в среднем составляет 7%.
Væringjavegr ᛝ Fennoskandia
Особенности Псковско-Новгородского ареала
Генетический анализ основного русскоязычного населения Восточно-Европейской равнины показывает, что основная часть этой популяции характеризуется низким уровнем генетической дифференциации как по мтДНК, так и по линиям Y-хромосомы. Генетические различия между популяциями исчезают, если их сгруппировать в соответствии с диалектным разнообразием русского языка и данными антропологии. Анализ популяций, представляющих этнографическую группу русскоязычных поморов, приводит к значительному увеличению значений межпопуляционной генетической дифференциации. Поморы отличаются высоким генетическим сходством с финно-угорским и, в некоторых случаях, со скандинавским населением Северной Европы, а также проявляют высокую степень многоступенчатости генофонда, вызванную влиянием генетического дрейфа.
Популяции объединяли в группы учитывая:
– (А) по говорам и наречиям русского языка: южное наречие (Белгород, Орёл, Калуга, Тула), северное наречие (Ярославль), среднерусские говоры (Великий Новгород, Волот, Псков, Владимир, Нижний Новгород);
– (Б) по диалектным зонам: юго-западная зона (Белгород, Орёл, Калуга, Тула), северо-западная зона (Великий Новгород, Волот, Псков), северо-восточная зона (Владимир, Нижний Новгород, Ярославль);
– (В) в соответствии с данными антропологии: предполагаемые потомки словен новгородских и псковских кривичей (Великий Новгород, Волот, Псков), кривичей (Владимир, Нижний Новгород, Ярославль), вятичей (Калуга, Тула), северян (Орёл, Белгород).
Анализ изменчивости нуклеотидных последовательностей гипервариабельного сегмента 1 (ГВС1) мтДНК в популяциях Тульской, Калужской, Владимирской, Ярославской, Псковской, Новгородской (Великий Новгород и Волот), Белгородской, Нижегородской, Орловской, Саратовской, Курской и Костромской областей и Ставропольского края (проанализировано 913 человек) – значение межпопуляционной дифференциации FST составляет 0,35% (P = 0,0007).
Анализ данных об изменчивости нуклеотидных последовательностей ГВС1 мтДНК в популяциях северо-западной зоны (Великий Новгород, Волот и Псков) был проведён в сравнении с другими славянскими популяциями Восточно-Европейской равнины (русские, поляки), а также популяциями соседних народов: балтскими (литовцы) и финскими (эстонцы, карелы и финны). Исследование показало, что степень дифференциации между проанализированными популяциями низка – FST = 0,38%, P = 0. Однако некоторые популяции демонстрируют высокое генетическое сходство.
Выделена отдельная группа населения, которая включает русскоязычные популяции северо-западной зоны (Великий Новгород и Псков), северо-восточную польскоязычную группу (Suwałki) и популяции балтийского и финского происхождения, что приводит к увеличению степени межгрупповой дифференциации (0,15%). Это свидетельствует о своеобразии генетической структуры этих популяций как по отношению к остальным славянским популяциям (русским и полякам; 0,52%), так и к популяциям балтийского и финского происхождения (0,76%). Анализ генофондов северо-западных русскоязычных и северо-восточных польскоязычных населений в сравнении с их непосредственными соседями – литовцами и эстонцами – показал значительные различия между ними (0,73%).
Таким образом, биологическое разнообразие является результатом формирования уникальной генетической композиции, сложившейся в результате взаимодействия трёх генофондов – славянского, балтского и финно-угорского. Анализ структуры митохондриального генофонда популяций Великого Новгорода, Пскова и Suwałki показал наличие только одной генетической компоненты – гаплогруппы U5a, которая распространена в этих популяциях с более высокой частотой (в среднем 16%), чем в соседних славянских, балтийских и финно-угорских популяциях, где её частота в среднем составляет 7%.
Væringjavegr ᛝ Fennoskandia
По трём генетическим системам были изучены народы, говорящие на языках балто-славянской лингвистической группы – шестнадцать народов по единой обширной панели маркеров:
балтские народы – латыши и литовцы;
восточные славяне – белорусы, русские, украинцы и русины;
западные славяне – кашубы, поляки, словаки, сорбы и чехи;
южные славяне – болгары, боснийцы, македонцы, сербы, словенцы и хорваты.
Восточные славяне – русские, белорусы, украинцы и русины – отчётливо группируются в единый генетический кластер. Исключение составляют русскоязычные популяции северного ареала, которые генетически отдалены от восточных славян и тяготеют к соседним финно-угорским и скандинавским популяциям.
Из западных славян: чехи, и в меньшей степени словаки, отличаются от восточных славян, и смещены в сторону германоязычных народов и других западноевропейских популяций. В то время как поляки наиболее близки к восточным славянам.
Южные славяне, формируют дисперсную группу, которая внутренне поделена на: западные (словенцы, хорваты и боснийцы) и восточные (македонцы и болгары) регионы, с сербами посередине. При этом, словенцы генетически близки к венграм, а восточная ветвь южных славян группируется с румынами и до некоторой степени с греками.
Балтские народы – латыши и литовцы – имеют генетическую близость к эстонцам и к западным беларусам. Оказалось также, что балтские популяции близки к волжской группе финно-угорских народов (особенно к эрзянам).
У балто-славянских популяций почти весь спектр представлен двумя основными цветами предковых компонентов: синий (k3) и голубой (k2). Предковый компонент k3 максимален у балтских популяций, превалирует у восточных славян (80-95%) и снижается у южных славян (55-70%). Напротив, k2 (голубой) более характерен для популяций средиземноморского и кавказского регионов и снижается к северу Европы. У южных славян на него приходится примерно 30%, у западных славян происходит снижение до 20%, а у северных русскоязычных и балтских популяций – до 5%.
У славянской этногруппы также присутствует лимонно-жёлтый компонент в предковом спектре, обозначаемый как k5, который значительно распространён только среди восточных славян, и в особенности среди северного кластера. По происхождению этот компонент является сибирским и составляет основную часть спектра для популяций Сибири. Тёмно-зелёный компонент k4 характерен для популяций Южной Азии, а также распространен на Ближнем Востоке и Средиземноморье и встречается низкой частотой у южных славян и других народов Балканского полуострова.
Популяционными генетиками было подсчитано число общих гаплотипов между «западно-восточными» славянами и восемью другими группами народов Европы:
1) южными славянами (болгары, боснийцы, македонцы, словенцы, хорваты);
2) популяциями Западной Европы (итальянцы, немцы, французы);
3) балтскими популяциями (латыши, литовцы);
4) популяциями северо-восточной Европы (вепсы, карелы, финны, эстонцы);
5) популяциями центральной Европы, ареал которых находится между западно-восточными и южными славянами – «между-славянские популяции»; разноязычные популяции: гагаузы, венгры, румыны);
6) греками;
7) популяциями волжского региона и Приуралья (башкиры, коми, эрзя, мокша, татары, удмурты, чуваши);
8) северокавказскими популяциями (адыги, балкарцы, ногайцы).
При введении условного «балто-славянского эталона» общих гаплотипов, часть окружающих неславянских популяций будет иметь меньшее родство с западно-восточными славянами, чем южные славяне – народы Поволжья, Западной Европы, Кавказа, а также греки. Родство генофондов балтов и популяций северо-восточной Европы в два раза выше (вепсы, карелы, финны, латыши, литовцы, поморы, эстонцы).
Согласно принципу эквидистантности, если система популяций развивается без больших внешних влияний, то постепенное разделение популяций приводит к линейному накоплению как лингвистического, так и генетического разнообразия. В результате генетическое разнообразие примерно одинаково на всех уровнях, будь то между популяциями одного народа, между этносами одной ветви или между их усредненными характеристиками. Величины генетического разнообразия славяноязычного населения на разных иерархических уровнях резко различаются – 0.06, 0.03, 0.01. Это указывает на активное смешение с окружающими народами. Наибольшая изменчивость приходится на самый древний уровень, что выявляет взаимодействия на ранних этапах истории славяноязычных популяций.
Тесты Y-хромосом, мтДНК и аутосомных маркеров показали чрезвычайно высокую корреляцию между генетикой и географическим положением популяций (0,80-0,95). Очень высокая корреляция также обнаружена между генетикой и лингвистикой (0,74-0,78). Однако, при исключении географического фактора, частная корреляция с лингвистикой становится намного ниже (0,3 для мтДНК и 0,2 для остальных двух систем), в то время как для всех трёх генетических систем корреляция с географией при исключении фактора лингвистики остается высокой (0,5 для мтДНК и 0,8 для остальных двух систем). Это указывает на то, что основная связь обусловлена географическим фактором, а высокая связь с лингвистикой объясняется тем, что народы, говорящие на родственных языках, часто являются и географическими соседями.
Генетики смогли выделить два основных субстрата, которые были ассимилированы славянами, впитав в себя местные популяции. «Центрально-восточноевропейский субстрат» был принят как западными, так и восточными славянами, с высокими частотами гаплогруппы R1a. Другой субстрат, «южно-восточноевропейский», был впитан южными славянами, с высокими частотами гаплогруппы I2a.
Особенности Псковско-Новгородского ареала
Генетический анализ основного русскоязычного населения Восточно-Европейской равнины показывает, что основная часть этой популяции характеризуется низким уровнем генетической дифференциации как по мтДНК, так и по линиям Y-хромосомы. Генетические различия между популяциями исчезают, если их сгруппировать в соответствии с диалектным разнообразием русского языка и данными антропологии. Анализ популяций, представляющих этнографическую группу русскоязычных поморов, приводит к значительному увеличению значений межпопуляционной генетической дифференциации. Поморы отличаются высоким генетическим сходством с финно-угорским и, в некоторых случаях, со скандинавским населением Северной Европы, а также проявляют высокую степень многоступенчатости генофонда, вызванную влиянием генетического дрейфа.
Популяции объединяли в группы учитывая:
– (А) по говорам и наречиям русского языка: южное наречие (Белгород, Орёл, Калуга, Тула), северное наречие (Ярославль), среднерусские говоры (Великий Новгород, Волот, Псков, Владимир, Нижний Новгород);
– (Б) по диалектным зонам: юго-западная зона (Белгород, Орёл, Калуга, Тула), северо-западная зона (Великий Новгород, Волот, Псков), северо-восточная зона (Владимир, Нижний Новгород, Ярославль);
– (В) в соответствии с данными антропологии: предполагаемые потомки словен новгородских и псковских кривичей (Великий Новгород, Волот, Псков), кривичей (Владимир, Нижний Новгород, Ярославль), вятичей (Калуга, Тула), северян (Орёл, Белгород).
Анализ изменчивости нуклеотидных последовательностей гипервариабельного сегмента 1 (ГВС1) мтДНК в популяциях Тульской, Калужской, Владимирской, Ярославской, Псковской, Новгородской (Великий Новгород и Волот), Белгородской, Нижегородской, Орловской, Саратовской, Курской и Костромской областей и Ставропольского края (проанализировано 913 человек) – значение межпопуляционной дифференциации FST составляет 0,35% (P = 0,0007).
Анализ данных об изменчивости нуклеотидных последовательностей ГВС1 мтДНК в популяциях северо-западной зоны (Великий Новгород, Волот и Псков) был проведён в сравнении с другими славянскими популяциями Восточно-Европейской равнины (русские, поляки), а также популяциями соседних народов: балтскими (литовцы) и финскими (эстонцы, карелы и финны). Исследование показало, что степень дифференциации между проанализированными популяциями низка – FST = 0,38%, P = 0. Однако некоторые популяции демонстрируют высокое генетическое сходство.
Выделена отдельная группа населения, которая включает русскоязычные популяции северо-западной зоны (Великий Новгород и Псков), северо-восточную польскоязычную группу (Suwałki) и популяции балтийского и финского происхождения, что приводит к увеличению степени межгрупповой дифференциации (0,15%). Это свидетельствует о своеобразии генетической структуры этих популяций как по отношению к остальным славянским популяциям (русским и полякам; 0,52%), так и к популяциям балтийского и финского происхождения (0,76%). Анализ генофондов северо-западных русскоязычных и северо-восточных польскоязычных населений в сравнении с их непосредственными соседями – литовцами и эстонцами – показал значительные различия между ними (0,73%).
Таким образом, биологическое разнообразие является результатом формирования уникальной генетической композиции, сложившейся в результате взаимодействия трёх генофондов – славянского, балтского и финно-угорского. Анализ структуры митохондриального генофонда популяций Великого Новгорода, Пскова и Suwałki показал наличие только одной генетической компоненты – гаплогруппы U5a, которая распространена в этих популяциях с более высокой частотой (в среднем 16%), чем в соседних славянских, балтийских и финно-угорских популяциях, где её частота в среднем составляет 7%.
Væringjavegr ᛝ Fennoskandia